
Order-level phylogeny
In birds, higher-level clades are traditionally treated as orders, recognised by the ending -iformes. The number of orders, however, is not defined and may vary among authors. Here, I follow the AviList checklist v2025 in recognising 46 orders. (link)

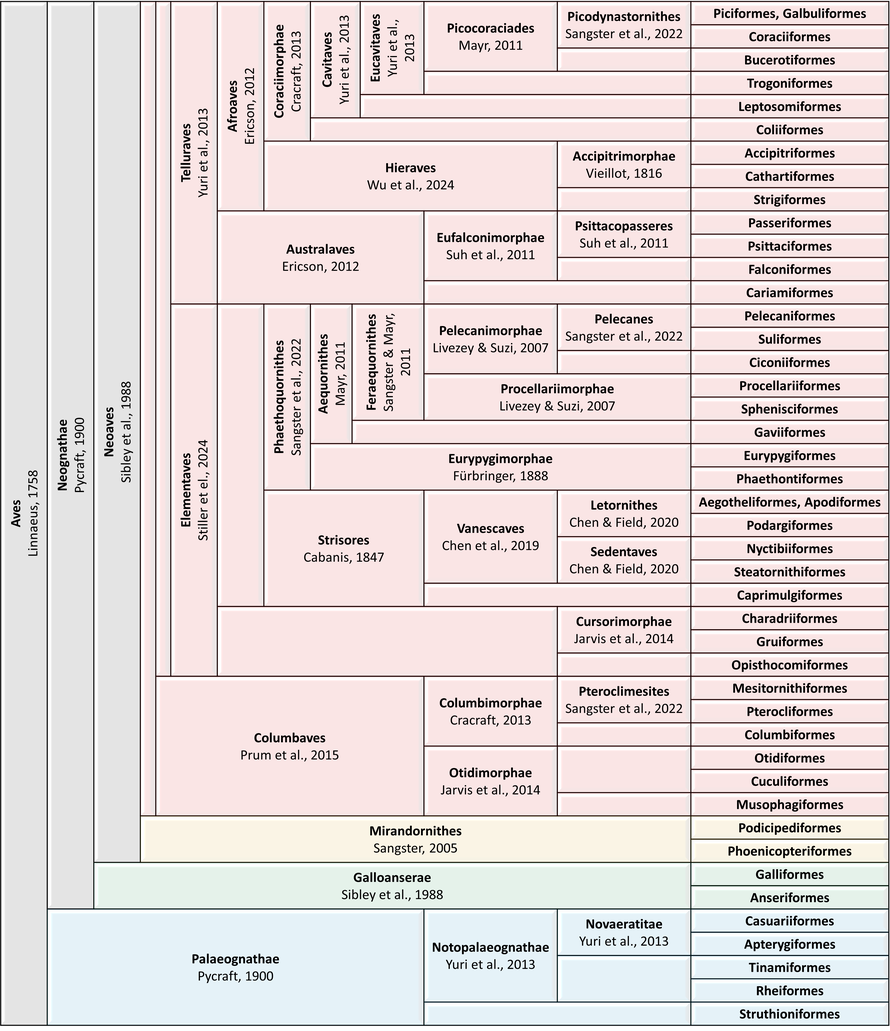
Traditional order-level classification of extant Aves according to Sangster et al. (2022), Wu et al. (2024),
Stiller et al. (2024), and AviList checklist v2025.
Bibliography
Braun EL, and Kimball RT (2021), Data types and the phylogeny of Neoaves, Birds 2, 1-22. (pdf)
Braun EL, Oliveros CH, White Carreiro ND, Zhao M, Glenn TC, Brumfield RT, Braun MJ, Kimball RT, and Faircloth BC (2024), Testing the mettle of METAL: A comparison of phylogenomic methods using a challenging but well-resolved phylogeny, bioRxiv [preprint] (pdf)
Bravo GA, Schmitt CJ, and Edwards SV (2021), What have we learned from the first 500 avian genomes? Annu. Rev. Ecol. Evol. Syst. 52, 611-639. (abstract)
Brocklehurst N, and Field DJ (2024), Tip dating and Bayes factors provide insight into the divergences of crown bird clades across the end-Cretaceous mass extinction, Proc. R. Soc. B 291, e:20232618. (pdf)
Burleigh JG, Kimball RT, and Braun EL (2015), Building the avian tree of life using a large-scale, sparse supermatrix, Mol. Phylogenet. Evol. 84, 53-63. (abstract)
Chen A, and Field DJ (2020), Phylogenetic definitions for Caprimulgimorphae (Aves) and major constituent clades under the International Code of Phylogenetic Nomenclature, Vertebr. Zool. 70, 571-585. (pdf)
Claramunt S, and Cracraft J (2015), A new time tree reveals Earth history´s imprint on the evolution of modern birds, Sci. Adv. 1, e:1501005. (pdf)
Claramunt S, Braun EL, Cracraft J, Fjeldså J, Ho SYW, Houde P, Nguyen JMT, and Stiller J (2024), Calibrating the genomic clock of modern birds using fossils, Proc. Natl. Acad. Sci. U.S.A. 121, e:2405887l21. (free pdf)
Claramunt S, Sheard C, Brown JW, Cortés-Ramirez G, Cracraft J, Su MM, Weeks BC, and Tobias JA (2025), A new time tree of birds reveals the interplay between dispersal, geographic range size, and diversification, Curr. Biol. (pdf)
Cloutier A, Sackton TB, Grayson P, Clamp M, Baker WJ, and Edwards SV (2019) Whole-genome analyses resolve the phylogeny of flightless birds (Palaeognathae) in the presence of an empirical anomaly zone, Syst. Biol. 68, 937-955. (free pdf)
Cracraft J (2013), Avian higher-level relationships and classification: Vol.1, Non-passerines, In: The Howard and Moore Complete Checklist of the Birds of the World (Dickinson EC, and Remsen JV, eds.), 4th edition, xxi-xliii, Aves Press, Eastbourne, UK. (link)
Cracraft J., Houde
P, Ho SYW, Mindell DP, Fjeldså J, Lindow B, Edwards SV, Rahbek C, Mirarab S, Warnow T, Gilbert MTP, Zhang G, Braun EL, and Jarvis ED (2015), Response to comment on “Whole-genome analyses resolve early branches in the tree of life of modern birds“, Science 349, 6255. (abstract)
Crouch NMA, Ramanauskas K, and Igic B (2019), Tip-dating and the origin of Telluraves, Mol. Phylogenet. Evol. 131, 55-63. (abstract)
de Quieroz K, Cantino PD, and Gauthier JA (Eds.) (2020), Phylonyms: a companion to the PhyloCode, CRC Press, Boca Raton, FL, USA. (link)
del Hoyo J, Collar NJ, Christie DA, Elliott A, and Fishpool LDC (2014), Illustrated Checklist of the Birds of the World, volume I (Non-passerines), published by Lynx Edicions in association with BirdLife. (link)
Ericson PGP, Anderson CL, Britton T, Elzanowski A, Johansson US, Kallersjo M, Ohlson JI, Parsons TJ, Zuccon D, and Mayr G (2006), Diversification of Neoaves: integration of molecular sequence data and fossils, Biol. Lett. 2, 543-547. (abstract)
Ericson PGP (2012), Evolution of terrestrial birds in three continents: biogeography and parallel radiations, J. Biogeogr. 39, 813-824. (abstract)
Gatesy J, and Springer MS (2022), Phylogenomic coalescent analyses of avian retroelements infer zero-length branches at the base of Neoaves, emergent support for controversial clades, and ancient introgressive hybridization in Afroaves, Genes 13, e:1167. (free pdf)
Gilbert PS, Wu J, Simon MW, Sinsheimer JS, and Alfaro ME (2019), Filtering nucleotide sites by phylogenetic signal to noise ratio increases confidence in the Neoaves phylogeny generated from ultraconserved elements, Mol. Phylogenet. Evol. 126, 116-128. (abstract)
Hackett SJ, Kimball RT, Reddy S, Bowie RCK, Braun EL, Braun MJ, Chojnowski JL, Cox WA, Han KL, Harshman J, Huddleston CJ, Marks BD, Miglia KJ, Moore WS, Sheldon FH, Steadman DW, Witt CC, and Yuri T (2008), A phylogenetic study of birds reveals their evolutionary history, Science 320, 1763-67. (abstract)
Harvey MG, Bravo GA, Claramunt S, Cuervo AM, Derryberry GE, Battilana J, Seeholzer GF, McKay JS, O'Meara BC, Faircloth BC, Edwards SV, Perez-Eman J, Moyle RG, Sheldon FH, Aleixo A, Smith BT, Chesser RT, Silveira LF, Cracraft J, Brumfield RT, and Derryberry EP (2020), The evolution of a tropical biodiversity hotspot, Science 370, 1343-48. (pdf) (hi-res tree) [note: focussing on the suboscine passerines, i.e. Eurylaimides and Tyrannides]
Heine C, Zoethout J, and Müller RD (2013), Kinematics of the South Atlantic rift, Solid Earth 4, 215-253. (pdf)
Houde P, Braun EL, Narula N, Minjares U, and Mirarab S (2019), Phylogenetic signal of indels and the Neoavian radiation, Diversity 11, e:108. (free pdf)
Jarvis ED, Mirarab S, Aberer AJ, Li B, Houde P, Li C, Ho SYW, Faircloth BC, Nabholz B, Howard JT, Suh A, Weber CC, de Fonseca RP, Li J, Zhang F, Li H, Zhou L, Narula N, Liu L, Ganapathy G, Boussau B, Bayzid MS, Zavidovych V, Subramanian S, Gabaldón T, Capella-Gutiérrez S, Huerta-Cepas J, Rekepalli B, Munch K, Schierup M, Lindow B, Warren WC, Ray D, Green RE, Bruford MW, Zhan X, Dixon A, Li S, Li N, Huang Y, Derryberry EP, Bertelsen MF, Sheldon FH, Brumfield RT, Mello CV, Lovell PV, Wirthlin M, Cruz Schneider MP, Prosdocimi F, Samaniego JA, Vargas Velazquez AM, Alfaro-Núñez A, Campos PF, Petersen B, Sicheritz-Ponten T, Pas A, Bailey T, Scofield P, Bunce M, Lambert DM, Zhou Q, Perelman P, Driskell AC, Shapiro B, Xiong Z, Zeng Y, Liu S, Li Z, Liu B, Wu K, Xiao J, Yinqi X, Zheng Q, Zhang Y, Yang H, Wang J, Smeds L, Rheindt FE, Braun M, Fjeldsa J, Orlando L, Barker FK, Jønsson KA, Johnson W, Koepfli KP, O´Brian S, Haussler D, Ryder OA, Rahbek C, Willerslev E, Graves GR, Glenn TC, McCormack J, Burt D, Ellegren H, Alström P, Edwards SV, Stamatakis A, Mindell DP, Cracraft J, Braun EL, Warnow T, Jun W, Gilbert MTP, and Zhang G (2014), Whole-genome analyses resolve early branches in the tree of life of modern birds, Science 346, 1320-31. (abstract)
Jetz W, Thomas GH, Joy JB, Hartmann K, and Mooers AO (2012), The global diversity of birds in space and time, Nature 491, 444-448. (abstract)
Kimball RT, Oliveros CH, Wang N, White ND, Barker FK, Field DJ, Ksepka DT, Chesser RT, Moyle RG, Braun MJ, Brumfield RT, Faircloth BC, Smith BT, and Braun EL (2019), A phylogenomic supertree of birds, Diversity 11, e:109. (free pdf)
Ksepka DT, Stidham TA, and Williamson TE (2017), Early Paleocene landbird supports rapid phylogenetic and morphological diversification of crown birds after the K-Pg mass extinction, Proc. Natl. Acad. Sci. U.S.A. 114, 8047-52. (pdf)
Kuhl H, Frankl-Vilches C, Bakker A, Mayr G, Nikolaus G, Boerno ST, Klages S, Timmermann B, and Gahr M (2021), An unbiased molecular approach using 3'UTRs resolves the avian family-level tree of life, Mol. Biol. Evol. 38, 108-127. (free pdf)
Linnaeus C (1758), Systema Naturæ, 10th edition, (link)
Livezey BC, and Zusi Rl (2007), Higher-order phylogeny of modern birds (Theropoda, Aves: Neornithes) based on comparative anatomy. II. Analysis and discussion, Zool. J. Linn. Soc. 149, 1-95. (pdf)
McCormack JE, Harvey MG, Faircloth BC, Crawford NG, Glenn TC, and Brumfield RT (2013), A phylogeny of birds based on over 1,500 loci collected by target enrichment and high-throughput sequencing, PLOS ONE 8, e:54848. (pdf)
McTavish EJ, Gerbracht JA, Holder MT, Iliff MJ, Lepage D, Rasmussen PC, Redelings BD, Sanchéz Reyes LL, and Miller ET (2025), A complete and dynamic tree of birds, Proc. Natl. Acad. Sci. U.S.A. 122, e:2409658122. (pdf)
Mirarab S, Rivas-González I, Feng S, Stiller J, Fang Q, Mai U, Hickey G, Chen G, Brajuka N, Fedrigo O, Formenti G, Wolf JBW, Howe K, Antunes A, Schierup MH, Paten B, Jarvis ED, Zhang G, and Braun EL (2024), A region of suppressed recombination misleads neoavian phylogenomics, Proc. Natl. Acad. Sci. U.S.A. 121, e:23l9506121. (pdf)
Mitchell KJ, Cooper A, and Phillips MJ (2015), Comment on "Whole-genome analyses resolve early branches in the tree of life of modern birds", Science 351, 1460. (pdf)
Prum RO, Berv JS, Dornburg A, Field DJ, Townsend JP, Lemmon EM, and Lemmon AR (2015), A comprehensive phylogeny of birds (Aves) using targeted next-generation DNA sequencing, Nature 526, 569-57. (abstract)
Pycraft WP (1900), On the morphology and phylogeny of the Palaeognathae (Ratitae and Crypturi) snd Neognathae (Carinatae), Trans. Zool. Soc. Lond. 15, 149-290. (abstract)
Reddy S, Kimball RT, Pandey A, Hosner PA, Braun MJ, Hackett SJ, Han KL, Harshman J, Huddleston CJ, Kingston S, Marks BD, Miglia KJ, Moore WS, Sheldon FH, Witt CC, Yuri T, and Braun EL (2017), Why do phylogenomic data sets yield conflicting trees? Data type influences the avian tree of life more than taxon sampling, Syst. Biol. 66, 857-879. (abstract)
Rheindt FE, Donald PF, Donsker DB, Gerbracht JA, Iliff MJ, Lepage D, Norman JA, Rasmussen PC, Schodde R, Schulenberg TS, Areta JI, Brammer FP, Chesser RT, Dowsett RJ, Peterson A, Alström P, Stervander M, Remsen JV, Garnett ST, Homberger DG, Lei F, and Christidis L (2025), AviList: a unified global bird checklist, Biodivers. Conserv. (free pdf)
Sangster G (2005a), A name for the flamingo-grebe clade, Ibis 147, 612-615. (abstract)
Sangster G (2005b), A name for the clade formed by owlet-nightjars, swifts and hummingbirds (Aves), Zootaxa 799, 1-6. (pdf)
Sangster G, and Mayr G (2021), Feraequornithes; a name for the clade formed by Procellariiformes, Sphenisciformes, Ciconiiformes, Suliformes and Pelecaniformes (Aves), Vertebr. Zool. 71, 49-53. (free pdf)
Sangster G, Braun EL, Johansson US, Kimball RT, Mayr G, and Suh A (2022), Phylogenetic definitions for 25 higher-level clade names of birds, Avian Res. e:100027. (free reading)
Sayol F, Steinbauer
MJ, Blackburn TM, Antonelli A, and Faurby S (2020), Anthropogenic extinctions conceal widespread flightlessness in birds. Sci. Adv. 6, eabb6095. (pdf)
Sibley CG, Ahlquist JE, and Monroe BL (1988), A classification of the living birds of the world based on DNA-DNA hybridization studies, Auk 105, 409-423. (pdf)
Springer MS, and Gatesy J (2024), A new phylogeny for Aves is compromised by pervasive misalignment and homology problems, Proc. Natl. Acad. Sci. U.S.A. 121, e:2406494l21. (pdf)
Stiller J (2025), Synthesizing decades of research into one tree for birds, Proc. Natl. Acad. Sci. U.S.A. 122, e:2507805122. (pdf)
Stiller J, Feng S, Chowdhury AA, Rivas-González I, Duchêne DA, Fang Q, Deng Y, Kozlov A, Stamatakis A, Claramunt S, Nguyen JMT, Ho SYW, Faircloth BC, Haag J, Houde P, Cracraft J, Balaban M, Mai U, Chen G, Gao R, Zhou C, Xie Y, Huang Z, Cao Z, Yan Z, Ogilvie HA, Nakhleh L, Lindow B, Morel B, Fjeldså J, Hosner PA, da Fonseca RR, Petersen B, Tobias JA, Székely T, Kennedy JD, Reeve AH, Liker A, Stervander M, Antunes A, Tietze DT, Bertelsen M, Lei F, Rahbek C, Graves GR, Schierup MH, Warnow T, Braun EL, Gilbert MTP, Jarvis ED, Mirarab S, and Zhang G (2024), Complexity of avian evolution revealed by family-level genomes, Nature 629, 851-860. (pdf)
Suh A, Paus M, Kiefmann M, Churakov G, Franke FA, Brosius J, Kriegs JO, and Schmitz J (2011), Mesozoic retroposons reveal parrots as the closest living relatives of passerine birds, Nat. Commun. 2, e:443. (pdf)
Suh A, Smeds L, and Ellegren H (2015), The dynamics of incomplete lineage sorting across the ancient adaptive radiation of neoavian birds, PLOS Biology 13. (pdf)
Suh A (2016), The phylogenetic forest of bird trees contains a hard polytomy at the root of Neoaves, Zool. Scr. 45, 50-62. (pdf)
Tamashiro RA, White ND, Braun MJ, Faircloth BC, Braun EL, and Kimball RT (2019), What are the roles of taxon sampling and model fit in tests of cyto-nuclear discordance using avian mitogenomic data?, Mol. Phylogenet. Evol. 130, 132-142. (abstract)
Vellekoop J, Sluijs A, Smit J, Schouten S, Weijers JWH, Sinninghe Dámste JS, and Brinkhuis H (2014), Rapid short-term cooling following the Chicxulub impact at the Cretaceous–Paleogene boundary, Proc. Natl. Acad. Sci. U.S.A. 111, 7537-41. (pdf)
Vieillot LP (1816), Analyse d’une nouvelle ornithologie élémentaire, pp. 1-70, Paris. (link)
Wang N, Braun EL, Liang B, Cracraft J, and Smith SA (2021), Categorical edge-based analyses of phylogenomic data reveal conflicting signals for difficult relationships in the avian tree, Mol. Phylogenet. Evol. 174, e:107550. (abstract)
Wu S, Rheindt FE, Zhang J, Wang J, Zhang L, Quan C, Li Z, Wang M, Wu F, Qu Y, Edwards SV, Zhou Z, and Liu L (2024), Genomes, fossils, and the concurrent rise of modern birds and flowering plants in the Late Cretaceous, Proc. Natl. Acad. Sci. U.S.A. 121, e:23l9696121. (pdf)
Wu S, Rheindt FE, Zhang J, Wang J, Zhang L, Quan C, Li Z, Wang M, Wu F, Qu Y, Edwards SV, Zhou Z, and Liu L (2024), Reply to Claramunt et al.: Robustness of the Cretaceous radiation of crown aves, Proc. Natl. Acad. Sci. U.S.A. 121, e:24l2448121. (pdf)
Wu S, Rheindt FE, Zhang J, Wang J, Zhang I, Quan C, Li Z, Wang M, Wu F, Qu Y, Edwards SV, Zhou Z, and Liu L (2024), Reply to Springer and Gatesy: The impact of long branches and misalignments on phylogenetic analysis is minimal, Proc. Natl. Acad. Sci. U.S.A. 121, e:2409344l21. (pdf)
Wu S, Tao Z, Liu L, Marshall CR, Edwards SV, Zhou Z, and Rheindt FE (2025), New fossils imply a deeper origin of modern birds in the Mesozoic, Natl. Sci. Rev. 12, e:nwaf238. (free pdf)
Yuri T, Kimball RT, Harshman J, Bowie RCK, Braun MJ, Chojnowski JL, Han KL, Hackett SJ, Huddleston CJ, Moore WS, Reddy S, Sheldon FH, Steadman DW, Witt CC, and Braun EL (2013), Parsimony and model-based analyses of indels in avian nuclear genes reveal congruent and incongruent phylogenetic signals, Biology 2, 419-444. (free pdf)
Zhao M, Thom G, Faircloth BC, Andersen MJ, Barker FK, Benz BW, Braun MJ, Bravo GA, Brumfield RT, Chesser RT, Derryberry EP, Glenn TC, Harvey MG, Hosner PA, Imfeld TS, Joseph L, Manthey JD, McCormack JE, McCullough JM, Moyle RG, Oliveros CH, White Carreiro ND, Winker K, Field DJ, Ksepka DT, Braun EL, Kimball RT, and Smith BT (2025), Efficient inference of macrophylogenies: Insights from the Avian Tree of Life, bioRxiv preprint server (pdf)